Notes For All Chapters Biology Class 11 CBSE
All animals including human beings depend on plants for their food. Green plant synthesize the food they need, and for all other organisms depend on them for their needs. Green plants carry out photosynthesis, a physio-chemical processor which they use the light energy to derive the synthesis of organic compounds.
Topic 1 Introduction to Photosynthesis
The energy required by all living organisms comes directly or indirectly from the sunlight.
Thus, sunlight plays an important role in fixation of CO2 through which, conversion of solar energy into chemical energy takes place. Water plays a significant role during this process.
Hence, photosynthesis is the process by which plants, some bacteria and some protistans uses the energy from sunlight to produce sugar, which through cellular respiration produce ATP, the fuel used by all living organisms.
Photosynthesis is an important phenomenon due to the following two reasons
(i) It is the primary source of all food on the earth.
(ii) It is also responsible for the release of oxygen into the atmosphere by green plants.
Photosynthesis is the only phenomenon of biological importance that can harvest the energy of sunlight.
Requirements of Photosynthesis
On the basis of outline knowledge about the role of light, green plants, CO2 etc in the process of photosynthesis, several simple experiments might be performed indicating that chlorophyll, light and carbon dioxide are essential components for photosynthesis to take place.
Necessity of Chlorophyll (Green Pigment of Leaf)
To start with this experiment two leaves are taken, one is a variegated leaf or a leaf that must partially covered with black paper and another leaf that must be exposed to light. When these leaves are tested for presence of starch, it was observed that photosynthesis had occurred only in the green parts of the leaves in the presence of light, which ensures that chlorophyll is essential for photosynthesis.
Necessity of Carbon Dioxide
To initiate the half leaf experiment (given by Moll), a part of leaf is enclosed in a test tube. The test tube contains some cotton soaked in KOH (KOH is used because it absorbs carbon dioxide) and the another half of leaf is exposed to light.
The set up is then allowed to stand in light for about few hours, when starch test was done, it was observed that the exposed part of leaf was tested positive for starch while, the portion that was enclosed in the tube tested negative. This indicates that, the CO2 is also essential for photosynthesis to take place.
The difference between the two parts of the leaves in the Moll’s half leaf experiment was due to the difference in the availability of CO2 outside and its absence inside the bottle.
Necessity of Light
The rate of photosynthesis is directly proportional to the intensity of light. The necessity of light for photosynthesis can be shown by fixing a dark paper on leaf of well watered, but destarched plant (plant can be destarched, by placing it in dark for 48 hours).
After fixing paper, the set-up is exposed to sunlight for 2-6 hrs. After removing paper, the test for starch is performed over leaf. Starch is produced only in area that received light, shows that light is necessary for photosynthesis.
Necessity of Water
Through radio labelling of oxygen in water molecule, it is confirmed that the 02 released during photosynthesis comes from H2O, not CO2.
Early Experiments
Study of photosynthesis started about hundreds of years ago. Prior to that researchers used to believe that plants gets all of their nourishment from soil only by the means of roots. Thus, several simple experiments led to the development of understanding of the process involved. Some early experiments performed by different scholars are as follows.
Joseph Priestley
He performed a series of experiments in 1770s that revealed about the essentiality of air in the growth of green plants. He observed that a burning candle or a respiring mouse in a closed space, (i. e., bell jar) soon gets extinguished and died due to suffocation respectively, because burning candle and animal that breathe the air soon get damages.
On the other hand after placing a mint plant in the bell jar along the burning candle and mouse, he observed that the mouse stayed alive and also the candle continue to burn for a longer time.
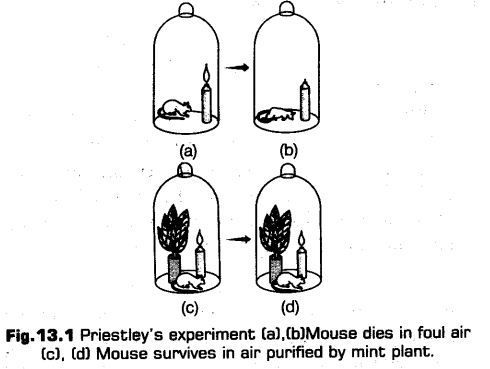
Conclusion
Priestley hypothesized that the foul air produced by the burning of candles and mouse respiration could be converted into pure air by plant (mint plant in this case).
Jan Ingenhousz (1730-1799)
He performed his experiments by using same set-up used by Priesdey. In his experiment with an aquatic plant, he showed that in bright sunlight, formation of small bubbles take place around the green parts, while in the dark, formation of those bubbles did not take place.
He did this by placing the experimental set-up once in the dark and once in the sunlight. The bubbles that he observed were of oxygen and showed that only green parts of plants could release oxygen.
Conclusion
He thus, concluded that sunlight is essential for the plant that purifies the foul air produced by burning candles or by breathing of animals.
Julius Von Sachs
He with his experiments in 1854 provided evidence that glucose is produced when plants grow, which is usually stored as starch. He later showed that a green substance, i. e., chlorophyll is found to be located in special bodies called chloroplast in plant cells.
Conclusion
He came to the conclusion that green parts are the place in the plants where production of glucose takes place and the same is stored in the form of starch.
The Engelmann (1843-1909)
He determined the action spectrum of photosynthesis by performing the interesting experiment with the help of a green alga, Cladophora. He splits light into its spectral components by using prism. He then illuminated the alga placed in a suspension of aerobic bacteria.
The bacteria were used to detect the sites of O2 evolution. On doing so, he noticed that the accumulation of bacteria was mainly in the region of the blue and red light of the split spectrum.
Conclusion
By the work done by him at first, action spectrum of photosynthesis was thus described, which roughly resembles the absorption spectra of chlorophyll-a and b.
Therefore, the key features of the process of photosynthesis were known by the middle of the nineteenth century, which detailed that plants acquire light energy harvested from sunlight for the formation of carbohydrates (food) from CO2 and water.
The empirical equation thus, determined the totalprocess of photosynthesisfor organisms that evolves oxygen is understood as
![]()
Cornelius Van Neil (1897-1985)
He was a microbiologist, who made a significant contribution on the basis of his studies of purple and green bacteria (photosynthetic bacteria) in understanding the photosynthesis. He demonstrated that during the process of photosynthesis, the hydrogen from a suitable oxidisable compound transferred, which reduces CO2 to carbohydrates in the presence of sunlight.
![]()
With the help of this, he reaches to the conclusion that, photosynthesis is a light dependent phenomenon.
Further, he states that in photosynthetic bacteriaH2S acts as a hydrogen donor, which gets oxidised to sulphur, i.e., they do not evolve O2 during the process of photosynthesis. While in case of green plants H2O acts as a hydrogen donor, which evolves O2 as its oxidation product.
Thus, he inferred that O2, which is evolved by the green plants comes from the water (H2O) not from the CO2 N (later proved by the use of radioisotopic techniques).
Thus, the overall reaction of the photosynthesis is represented as
![]()
It is also to be noted that it is not a single step reaction that determines the process of photosynthesis instead it is a multi step process.
Conclusion
He concluded that the light is necessary to oxidize the photosynthetic substrate (H2S – Photosynthetic bacteria) and (H2O-Green plants) and release byproduct sulphur and 02 in Bacteria and Green plants respectively.
In absence of sunlight, this process does not occur.
Chloroplasts: The Site of Photosynthesis
These are the green plastids that function as the site of photosynthesis, i.e., helps in the synthesis of organic food.
The process of photosynthesis takes place in the green leaves of the plants because chloroplasts are abundantly present in the mesophyll cell of the leaves.
Chloroplast aligned themselves with their flat surfaces parallel to the walls of the mesophyll cells under optimum light intensities and they would be perpendicular to the walls of the mesophyll cells when the intensity goes very high.
Chloroplast is double membranous, DNA containing cell organalle.
Internally, a chloroplast contains a proteinaceous matrix or fluid called stroma, the membrane system called lamellae or thylakoids. At some places, the thylakoid gets aggregated to form stacks of discs, called
grana.
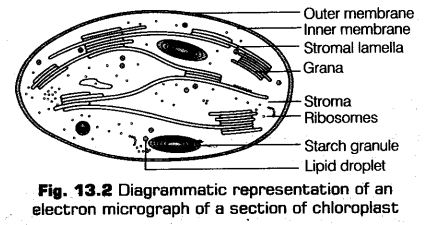
The clear division of labour occurs within the chloroplasts, i.e., the membrane system is responsible for the synthesis of ATP and NADPH (photochemical phase), whereas the stroma has enzymes, which are responsible for the reduction of carbon dioxide into carbohydrates and formation of sugars.
As the former sets of reactions are dependent on light so are called light reactions while, the latter is dependent on the products of light reactions, i.e., ATP and NADPH (and independent from direct sunlight), thus are called dark reactions.
It is to be noted that dark reactions does not rely that they occur in darkness or that they are not light dependent.
Pigments Involved in Photosynthesis
The pigments involved in the process of photosynthesis are called photosynthetic pigments. These pigments provides different shades of green in the leaves in different plants or in the leaves of same plant.
These pigments can easily be separated out by chromatographic technique (paper chromatography). On the basis of their significance, the photosynthetic pigments are of two types
(i) Primary pigments The pigment forms the main molecule of photosystem, e.g., Chlorophyll- a, b.
(ii) Accessory pigments. These support the function of primary pigments, e.g., Xanthophylls and carotenoids.
A cinematographic separation of the leaf pigments shows that it is not only the single pigment, which is responsible for the colour in the leaves. Instead the different shades in leaf are due to four different pigments that have different ability to absorb light at specific wavelength.
Different pigments present in leaf described below
(a) Chlorophyll-a (C55H72O5N4Mg) This is bright or blue green in the chromatogram. It is known to be the chief plant pigment associated with photosynthesis.
(b) Chlorophyll-b (C55H70O6N4Mg) This is yellow green in colour.
(c) Xanthophylls This is yellow in colour. These pigments are oxidised carotenoids.
(d) Carotenoids This is yellow to yellow-orange in colour. They are also known as ‘antenna pigment’.
Chlorophyll is the most abundant plant pigment found if the plants in the world. It contains magnesium (Mg+2 ) metal as its constituents.
Absorption Spectrum
It is the curve that shows the amount of different wavelength of lights absorbed by a substance. The graph given below shows the ability of chlorophyll-a to absorb lights of different wavelength.
Chtorophyll-a shows the maximum absorption peak at 450 nm and also shows another peak at 650 nm.
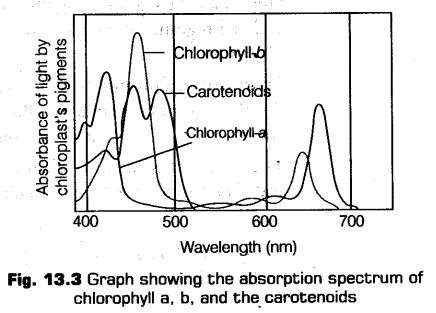
Absorption spectrum is constituted by the pigments like violet, blue, orange and red (400-500 and 600-700 nm).
The Emission spectrum is constituted by yellow and yellow-green pigment (500-600 nm).
Action Spectrum
It is the curve that depicts the relative rates of photosynthesis at different wavelengths of light. Now another graph given below, shows the wavelength at which maximum photosynthesis occurs at blue, violet and red wavelength in a plant (which is shown by chlorophyll-a).
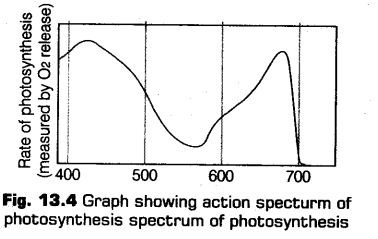
Hence, this concludes that chlorophyll-a is the chief pigment, which is majority responsible for the photosynthesis.The another graph given below show the action spectrum of photosynthesis, which coincide closely to the absorption spectrum of chlorophyll.
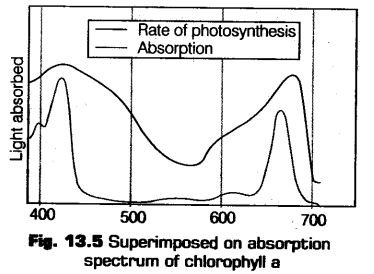
Hence, all three graphs together shows that the major part of the photosynthesis takes place in the blue and red regions, while some of the photosynthesis takes place at other wavelengths also in the visible spectrum.
Apart from chlorophyll-a (the major pigment,) which is mainly responsible for trapping of light, other thylakoid pigments such as chlorophyll-a, xanthophylls and carotenoids also absorb light, transferring energy to chlorophyll-a. These pigments are called accessory pigments.
These pigments enable a wider range of wavelength of incoming light to be utilised for photosynthesis and also provide protection to chlorophyll-a from photooxidation.
Photosynthetically Active Radiation (PAR)
The region of wavelength in which photosynthesis takes place normally. It ranges from 0.4μm to 0.7μm (400-700 nm).
Assimilatory Power
The chemicals (ATP and NADPH) synthesised in photosynthesis, termed as assimilatory power. They are used in the process to reduce CO2 into carbohydrates.
Topic 2 Mechanism of Photosynthesis
It was observed that the rate of photosynthesis is directly proportional to the intensity of light, i.e., rate increases with the increase in intensity of light till the plant achieved the saturation point.
The process ofphotosynthesis takes place in following two steps
(i) Light reaction or the photochemical phase.
(ii) Dark reaction or the biosynthetic phase.
Light Reaction (The Photochemical Phase)
Light reaction includes the following steps, i. e., absorption of light, splitting of water, release of oxygen and finally the formation of high energy chemical intermediates, i.e., ATP and NADPH. ‘
During the course of light reaction, light is trapped by photosynthetic pigments present in the quantasomes of grana thylakoids.
These photosynthetic pigments are organised into two discrete photochemical Light Harvesting Complexes (LHCs) known as Photosystem-I (PS-I) and Photosystem-II (PS-II).
Photosystems
The light harvesting complexes or photosystems are made up of hundreds of pigment molecules bounded by proteins. Each photosystem has a photocentre or reaction centre, where actual reaction takes place.
This reaction centre contains a special chlorophyll- molecule is fed by hundred other pigment molecules forms the light harvesting system called antennae. These antennae molecule absorb light of different wavelength, but shorter than reaction centre in order to make photosynthesis more efficient.
These photosystems are, named according to the sequence of their discoveries not in the sequence in which function during the light reaction.
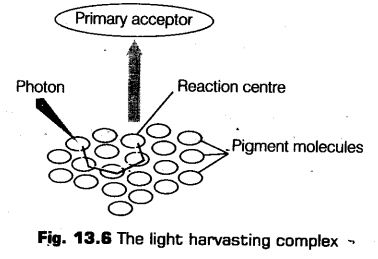 The reaction centre is different in both the photosystem as given below
The reaction centre is different in both the photosystem as given below
(i) In PS-I, the reaction centre or chlorophyll-a has peak of absorption at 700 nm, known as P700 .
(ii) In PS-II, the reaction centre has absorption peak at 680 nm hence, called P680 .
Electron Transport
The photosynthetic electron transport chain initiates by the absorbance of light by the photosystem-II. The red light of wavelength 680 nm is absorbed by the reaction centre of photosystem II due to which electrons becomes excited and jump into an orbit away from the atomic nucleus.
These electrons are then picked up by an electron acceptor, which passes them further to electron transport system consisting of cytochromes.
It is to be noted that this movement of electrons is down hill according to the redox potential scale (oxidation-reduction scale). The electrons of the electron transport chain are not used up in the chain instead they are further passed on to the pigments of PS-I.
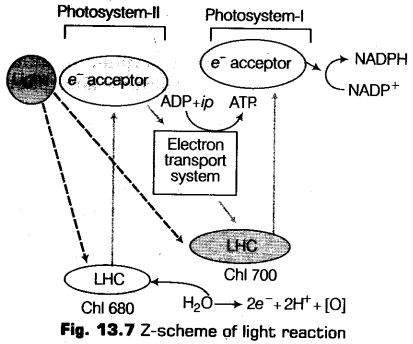
Now, like the PS-II, the electrons in the reaction centre of PS-I also gets excited on receiving red light of wavelength 700 nm and gets transferred to the another electron acceptor with hitter redox potential.
The electrons in this ease also moves downhill.
But this time the electrons does not move to a reaction centre or chlorophyll-a. Instead, moves to a molecule rich in energy and NADP+. On addition of these electrons the NADP+ gets reduced to NADPH + H+.
In 1960, Bendall and Hill discovered the Z-scheme of electron transport. It is a series of reactions that we have just studied above from the whole scheme of electron transfer initiating from PS-II, uphill to the acceptor molecule, down the electron transport chain to PS-I, excitation of electrons and then their transfer to another acceptor and finally downhill to NADP+ in order to get reduced to NADPH and H+.
Splitting of Water
The electrons are continuously supplied to the photosystems-II by the available electrons, which gets replaced due to the splitting of water. In this process the water splits into protons, electrons and oxygen. The complex for water splitting is associated with the photosystems-II that is located on the inner side of the thylakoid membrane.
These electrons thus, obtained by the splitting of water are needed to replace those electrons which are removed from the photosytems-I thus, are provided by photosystem-II.
2H2O —> 4H++O2 + 4e–
While all the electrons formed, are replaced, the protons gets accumulated in the lumen of the thylakoid and the oxygen gets into the atmosphere.
Photophosphorylation
Photophosphorylation is the process through which, ATP is synthesised from ADP and inorganic phosphate (P) by the cell organelles (like mitochondria and chloroplasts) with the help of energy from solar radiation.
Photophosphorylation in mitochondria is not light dependent, but it uses the energy by oxidation of nutrients to produce ATP, hence it is called oxidative phosphorylation.
The process of photophosphorylation is of two types
i. Non-cyclic Photophosphorylation
Non-cyclic photophosphorylation is type of photophosphorylarion in which both the photosystems (PS-I and PS-II) cooperate in light driven synthesis of ATP. During this cycle, the electron released from PS-II does not return to it hence, it is known as non-cyclic photophosphorylation. Both NADPH and ATP are formed during this reaction.
ii. Cyclic Photophosphorylation
It is the type of photophosphorylation in which only PS-I is taking part and the electron released from the reaction centre P700 returns to it after passing through a series of carrier i.e., circulation takes within the photosystem and the phosphorylation occurs due to cyclic flow of electrons.
iii. When non-eyeiie form of photophosphorylation gets stopped under certain conditions, the cyclic photophosphorylation occurs.
The cyclic photophosphorylation takes place in the stromal lamellae of the chloroplast. This happens because the stromal lamellae does not possess enzyme NADP reductase (essential for reducing NADP+ to NADPH) and PS-II. Thus, the excited electrons in the cyclic photophosphoiylation does not pass on to NADP+ instead it gets cycled back to the PS-I complex.
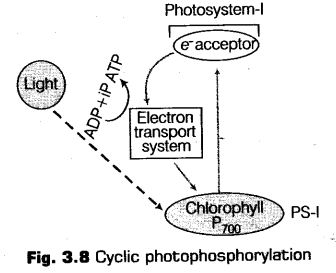
Hence, through the cyclic flow only the synthesis of ATP takes place.
Chemiosynthetic Hypothesis
This hypothesis was given by Peter Mitchell (1961) in order to explain the ATP synthesis in photosynthesis (also in respiration).
The synthesis of ATP is directly linked to the developinent of a proton gradient across the thylakoid membranes of a chloroplast.
The main difference that lies between the photosynthesis and respiration is the location where the accumulation of protons takes place. In chloroplast (photosynthesis), it occurs in thylakoid lumen while in mitochondria (respiration), it occurs in intermembrane space.
Now, the point arises that what causes the proton gradient across the membrane?
The development of proton gradient results due to the reasons given below
(i) As the water molecule splits into the inner side of the membrane the protons or hydrogen ions that are produced by the water splitting gets accumulate within the thylakoids lumen.
(ii) Transportation of protons takes place across the membrane when the electron moves through the photosystems. The primary acceptor of electron is located towards the outer side of the membrane, which transfers electron to the proton (H+) carrier and not to the electron carrier.
So, this molecule, while transporting an electron removes a proton from the stroma, thus, release of proton takes place into the inner side, i.e., on the lumen of the membrane.
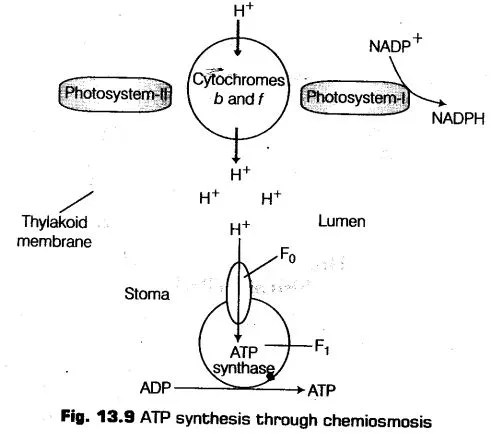
(iii) The enzyme NADP reductase is present on the stromal side of the membrane. Thus, alongwith the electrons that comes from the acceptor of electrons of PS-I, protons are also necessary to reduce NADP+ to NADPH + H+
Hence, protons in the stroma within the chloroplasts decreases in number, while accumulation of protons takes place in the lumen. Due to which the proton gradient is created across thylakoid membrane, which led to decrease in the pH at the side the lumen.
The gradient is broken down due to the movement of protons across the membrane to the stroma through the transmembrane channel of the F0 portion of the ATPase enzyme.
Therefore, the proton gradient is important as it is the gradient whose breakdown leads to the release of energy (ATP).
ATPase Enzyme
The enzyme ATPase consists of the following two parts
i. F0 Particle
This portion remains embedded in the membrane and forms a transmembrane channel, which carries out facilitated diffusion of protons across the membrane.
ii. F1 Particle
This portion protudes towards the outer surface of the thylakoid membrane which faces the stroma. Conformational change occurs in F1 particle of ATPase, which caused due to the breakdown of the gradient, which allows the enzyme to synthesise several molecules of ATP.
Thus, the chemiosmosis, for its functioning, requires a membrane, a proton pump, a proton gradient and ATPase enzyme. The ATP thus, produced will be used immediately in the biosynthetic reaction (in stroma), responsible for the fixing of CO2 and synthesis of sugar.
Dark Reaction (Biosynthetic Phase)
This phase does not require direct sunlight but it depends on the products of the light reaction, i.e., ATP and NADPH beside CO2 and water that drive the processes leading to the synthesis of food more accurately the sugars.
The O2 thus, produced in the light reaction of photosynthesis diffuses out of the chloroplast
As soon as the light becomes unavailable, the biosynthetic process continues for sometime and then eventually stops and starts again if the light is made available again.
Originally, this process is known as carbon-fixation or Photosynthetic Carbon Reduction (PCR) cycle.
These reactions are sensitive to temperature change, but are independent of light, hence called dark reaction. This takes place in the stroma of chloroplast.
Thus, assimilation of CO2 during photosynthesis is of two main types
i. C3 Pathway This pathway is followed by the plants when first product of CO2 fixation is a C3 acid, i.e., PGA.
ii. C4 Pathway This pathway is followed or shown by the plants in which first product ofCO2 fixation is a C4 acid, i. e., OAA.
Calvin Cycle (C3-Pathway)
This is a cycle biochemical pathway of reduction of CO2 or photosynthetic carbon, cycle, which was discovered by Calvin. The Calvin cycle runs in all photosynthetic plants, no matter they shows C3, C4 or any other pathways. It occurs in stroma of the chloroplast
Primary Acceptor of CO2 in C3 Pathway
After a long research and conducting many experiments it was concluded by the scientists that in C3 pathway, the acceptor molecule is a 5-carbon ketose sugar, i.e., Ribulose 5-phosphate (5 RuBP).
Calvin orC3 cycle has following three major steps

The steps mentioned above are the major steps. Another steps known as glycolytic reversal or formation of sugar takes place between reduction and regeneration.
1. Carboxylation
It is the most crucial step of the Calvin cycle in which utilisation of CO2 takes place for the carboxylation of RuBP.
Carboxylation is the process of fixation of CO2 into a stable organic intermediate.
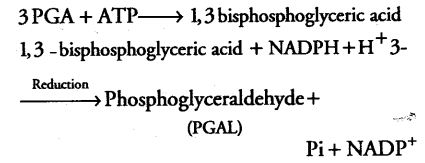
This reaction is catalysed by the enzyme RuBP carboxylase which finally results in the formation of two molecules of 3-PGA. As the RuBP carboxylase enzyme also has an activity of oxygenation. Thus, it more commonly known as RuBP carboxylase-oxygenase or RuBisCO.
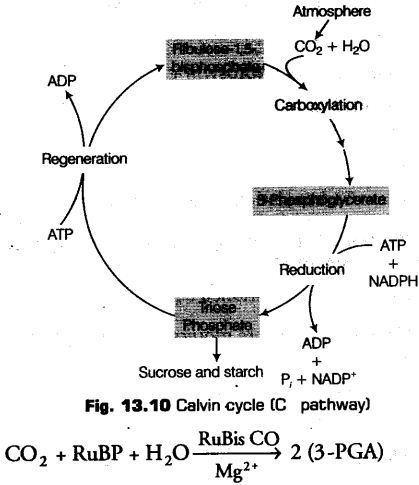
2. Reduction
After the carboxylation reaction, reduction of PGA takes place through a series of reactions leading to the formation of glucose. In this step, the ATP and NADPH (formed during the photochemical reaction) are utilised. It is to be noted that 2 molecules of ATP and 2 molecules of NADPH are utilised in this step for phosphorylation and for the reduction of CO2 respectively.
Hence, the fixation of 6 molecules of CO2 and 6 turns of the cycle are required in order to release one molecule of glucose from the pathway.
3. Regeneration
For the continuous and uninterrupted functioning of the Calvin cycle, there must be a regular supply of ATP, NADPH and also sufficient amount of RuBP is required. The regeneration of RuBP (CO2 acceptor) is a complex process and involves many types of sugar starting from triose (3C) to heptose (7C).
The regeneration step require one ATP molecule for phosphorylation. Hence, for every CO2 molecule that- enters the Calvin cycle, required are 3 molecules of ATP and 2 molecules of NADPH.
The cyclic phosphorylation takes place in order to meet the difference in the number of ATP and NADPH used in the dark reaction.
Thus, in order to produce one molecule of glucose through the Calvin pathway, 18 ATPs and 12 NADPHs are required.
This can he easily understood by the following table given below


Leave a Reply